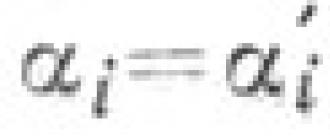Պասիվ և ակտիվ տրանսպորտ. Պասիվ և ակտիվ տրանսպորտ Նյութերի ակտիվ տեղափոխում
Ակտիվ փոխադրումը գործընթաց է, որի ընթացքում մոլեկուլը պետք է շարժվի մեմբրանի միջով՝ անկախ դրա կոնցենտրացիայի գրադիենտի ուղղությունից: Ամենից հաճախ դա տեղի է ունենում ավելի ցածր կոնցենտրացիա ունեցող տարածաշրջանից դեպի ավելի բարձր կոնցենտրացիա ունեցող տարածաշրջան և ուղեկցվում է ազատ էներգիայի աճով, որը կազմում է 5,71 logC2/C| կՋ-մոլ-1.
Ինչպես նշվեց ավելի վաղ, սա ավելի ցածր էլեկտրաքիմիական պոտենցիալ ունեցող վայրերից նյութերի ավելի մեծ արժեք ունեցող վայրեր տեղափոխելու գործընթաց է:
Քանի որ մեմբրանի մեջ ակտիվ փոխադրումը ուղեկցվում է Գիբսի էներգիայի ավելացմամբ, այն չի կարող տեղի ունենալ ինքնաբերաբար, այսինքն՝ նման գործընթացը պահանջում է զուգակցում որոշակի ինքնաբուխ ռեակցիայի հետ: Ընդհանուր առմամբ, դա կարող է իրականացվել երկու եղանակով. 2) միջնորդավորված մեմբրանի պոտենցիալով և/կամ իոնների կոնցենտրացիայի գրադիենտով` հատուկ փոխադրողների առկայության և թաղանթի առկայությամբ:
Առաջին դեպքում փոխադրումն իրականացվում է էլեկտրագենիկ իոնային պոմպերի միջոցով, որոնք սնվում են ATP հիդրոլիզի ազատ էներգիայով։ Դրանք դասակարգվում են որպես ինտեգրալ սպիտակուցների հատուկ համակարգեր և կոչվում են տրանսպորտային ATPases: Ներկայումս հայտնի են երեք տեսակի էլեկտրագենային իոնային պոմպեր, որոնք տեղափոխում են իոններ մեմբրանի միջով. K+ - Na+ - ATPase, յուրաքանչյուր ATP մոլեկուլի հիդրոլիզի ժամանակ թողարկված էներգիայի շնորհիվ կալիումի երկու իոն տեղափոխվում են բջիջ, իսկ երեք նատրիումի իոններ մղվում են: դուրս; Ca2+ - ATPase-ում, ATP հիդրոլիզի էներգիայի շնորհիվ, փոխանցվում են երկու կալցիումի իոններ. H+ պոմպում կա երկու պրոտոն։
Երկրորդ դեպքում նյութերի փոխադրումը երկրորդական է, որի համար խորապես ուսումնասիրվել են երեք սխեմաներ.
Իոնի միակողմանի փոխանցումը կոնկրետ կրիչի հետ համատեղ կոչվում է միապորտ: Այս դեպքում լիցքը մեմբրանի վրայով փոխանցվում է կամ կոմպլեքսով, եթե կրիչի մոլեկուլը էլեկտրականորեն չեզոք է, կամ դատարկ կրիչով, եթե փոխադրումն ապահովվում է լիցքավորված կրիչի կողմից: Փոխանցման արդյունքը կլինի իոնների կուտակումը մեմբրանի ներուժի նվազման պատճառով։ Այս ազդեցությունը նկատվում է, երբ կալիումի իոնները կուտակվում են էներգիա ունեցող միտոքոնդրիումներում վալինոմիցինի առկայության դեպքում:
Մեկ կրիչի մոլեկուլի մասնակցությամբ իոնների հակադարձ փոխանցումը կոչվում է հակապորտ։ Ենթադրվում է, որ փոխադրող մոլեկուլը փոխանցում է յուրաքանչյուր իոնով ուժեղ բարդույթ։ Փոխանցումը տեղի է ունենում երկու փուլով. նախ, մեկ իոն անցնում է թաղանթը ձախից աջ, այնուհետև երկրորդ իոնը հատում է թաղանթը: հակադարձ ուղղություն. Մեմբրանի ներուժը չի փոխվում: Ըստ երևույթին, այս գործընթացում շարժիչ ուժը փոխանցված իոններից մեկի կոնցենտրացիաների տարբերությունն է: Եթե ի սկզբանե տարբերություն չի եղել երկրորդ իոնի կոնցենտրացիայի մեջ, ապա փոխանցման արդյունքը կլինի երկրորդ իոնի կուտակումը առաջինի կոնցենտրացիաների տարբերության նվազման պատճառով։ Հակապորտի դասական օրինակ է կալիումի և ջրածնի իոնների փոխանցումը բջջային թաղանթով՝ հակաբիոտիկ նիգիրիցինի մասնակցությամբ։ Հարկ է նշել, որ կրող սպիտակուցների մեծ մասը գործում է որպես հակապորտ, այսինքն՝ նյութի շարժումը մեմբրանի միջով հնարավոր է դառնում միայն ինչ-որ բավականին կոնկրետ նյութի դիմաց, որն ունի նույն լիցքը, բայց շարժվում է հակառակ ուղղությամբ։
Այսպիսով, բջջի ցանկացած հիմնական բաղադրիչի արտազատումը կոնցենտրացիայի գրադիենտի երկայնքով կարող է վերահսկել հանդիպակաց նյութի շարժումը դեպի գրադիենտ և կատարել «աշխատանք», մինչև երկու շարժիչ ուժերը հավասարակշռվեն:
Նյութերի համատեղ միակողմանի փոխանցումը երկտեղանոց փոխադրողի մասնակցությամբ կոչվում է սիմպորտ։ Ենթադրվում է, որ թաղանթը կարող է պարունակել երկու էլեկտրականորեն չեզոք մասնիկներ՝ կրող կոմպլեքսում կատիոնով և անիոնով և դատարկ կրիչով։ Քանի որ նման տրանսպորտային համակարգում մեմբրանի ներուժը չի փոխվում, փոխադրումը կարող է առաջանալ իոններից մեկի կոնցենտրացիաների տարբերությամբ: Ենթադրվում է, որ սիմպորտի սխեմայի համաձայն, հետևում է, որ այս գործընթացը պետք է ուղեկցվի օսմոտիկ հավասարակշռության զգալի տեղաշարժով, քանի որ մեկ ցիկլում երկու մասնիկ տեղափոխվում են մեմբրանի միջով նույն ուղղությամբ:
Բավականին լավ զարգացած տեսությունների, իոնների փոխադրման մեխանիզմների և էնդոգեն առկայության պատճառով օրգանական նյութերխցում հնարավոր է դարձել մեկնաբանել դեղերի փորձարկումներից ստացված տվյալները (բաժին 6.3.3):
Համեմատությամբ Նկ. 6.10 ակտիվ տրանսպորտը կարող է ներկայացվել ինչպես ցույց է տրված Նկ. 6.11.
Այս դեպքում C կրիչը թաղանթի արտաքին կողմում կազմում է բարդ CA դեղամիջոցի հետ (L): Այն թափանցում է թաղանթը՝ մյուս կողմից պառակտելով L-ն։ Ակտիվ տրանսպորտի դեպքում մեմբրանի ներքին կողմում L-ի կոնցենտրացիան կարող է շատ ավելի մեծ լինել, քան արտաքին կողմի կոնցենտրացիան։ Ի տարբերություն պասիվ տրանսպորտի (նկ. 6.10), SA համալիրը, օգտագործելով ATP-ի էներգիան, վերածվում է SA համալիրի, որը հեշտությամբ բաժանվում է L-ի (Նկար 6.11 Հաշվի առնելով տրանսպորտի իրականացման համար էներգիայի ծախսերի անհրաժեշտությունը): SA-ի մեմբրանի հակառակ կողմում կարելի է ենթադրել, որ ներքին կողմում /(, (կտրվածքի հաստատուն) ավելի մեծ է, քան K0-ը: Սա թմրամիջոցների կրող համալիրի այսպես կոչված ասիմետրիկ ճեղքվածքն է:
Արտաքին ջրային փուլ
Համակենտրոնացում [L]0 Ակտիվություն (L)0
Կենդանի օրգանիզմներում ակտիվ փոխադրման մեխանիզմները լայն տարածում ունեն և կարելի է համարել բջջի հիմնարար գործառույթներից մեկը։ Օրինակ, բջիջներն ունեն կալիումի բարձր կոնցենտրացիա, իսկ նատրիումի ցածր կոնցենտրացիան՝ ի տարբերություն արտաբջջային տարածության, որտեղ այդ իոնները հակադարձ հարաբերությունների մեջ են։ Թաղանթները ազատորեն թափանցելի են երկու իոնների համար, և ասիմետրիկ բաշխումը պահպանվում է նատրիումը բջիջից անընդհատ «մղելով», իսկ կալիումը դեպի ներս: Ստամոքսում HC1-ի սեկրեցումը H+-ի և SG-ի իրական ակտիվ փոխադրումն է: Նմանատիպ մեխանիզմով յոդը կենտրոնանում է վահանաձև գեղձում։ Շաքարները տեղափոխվում են աղիներում և մոտակա երիկամային խողովակներում ավելի բարձր կոնցենտրացիաների դեմ: Ամինաթթուները նույն կերպ են վարվում աղիքներում, երիկամներում, մկաններում և ուղեղում: Երիկամային խողովակներով օրգանական թթուների (նապա-ամինոբենզոյան, հիպուրիկ) արտազատումը ակտիվ տրանսպորտային գործընթաց է։
Ակտիվ փոխադրման մեխանիզմը խիստ սպեցիֆիկ է, քանի որ այն ստեղծվել է բնության կողմից՝ բավարարելու օրգանիզմի կենսաբանական կարիքը հիմնական սննդանյութերի կամ դրանցից դրանց նյութափոխանակության արտադրանքները հանելու համար: Ինչ վերաբերում է ակտիվ տրանսպորտի ենթարկվող դեղերին, ապա այս դեպքում դրանք քիմիական կառուցվածքով պետք է մոտ լինեն մարմնի բնական նյութերին։ Պիրիմիդինի անալոգային ֆտորաֆուրը և երկաթը ներծծվում են աղիքներում ակտիվ տրանսպորտի միջոցով: Նույն մեխանիզմով լեվոդոպան ներթափանցում է արյունաուղեղային արգելքը։ Երիկամային խողովակները արտազատում են օրգանական թթուներին և հիմքերին պատկանող դեղամիջոցներ։
Ամփոփելով նյութերի տրանսմեմբրանային փոխադրման մեխանիզմների դիտարկումը, անհրաժեշտ է ևս մեկ անգամ ընդգծել, որ կյանքի ընթացքում բջջի սահմանները հատվում են տարբեր նյութերով, որոնց հոսքերը արդյունավետորեն կարգավորվում են: Այս խնդիրն իրականացվում է բջջային թաղանթով, որի մեջ ներկառուցված են տրանսպորտային համակարգեր, ներառյալ իոնային պոմպեր, կրող մոլեկուլների համակարգ և բարձր ընտրողական իոնային ալիքներ:
Առաջին հայացքից փոխանցման համակարգերի նման առատությունը անհարկի է թվում, քանի որ միայն իոնային պոմպերի շահագործումը հնարավորություն է տալիս ապահովել. բնութագրերըկենսաբանական տրանսպորտ՝ բարձր ընտրողականություն, նյութերի փոխանցում դիֆուզիոն ուժերի դեմ և էլեկտրական դաշտ. Այնուամենայնիվ, պարադոքսն այն է, որ կարգավորվող հոսքերի քանակը անսահման մեծ է, մինչդեռ կան ընդամենը երեք պոմպեր: Այս դեպքում հատուկ նշանակությունձեռք են բերում իոնային խոնարհման մեխանիզմներ, որոնք կոչվում են երկրորդական ակտիվ տրանսպորտ, որոնցում կարևոր դեր են խաղում դիֆուզիոն պրոցեսները։ Այսպիսով, նյութերի ակտիվ փոխադրման համադրությունը բջջաթաղանթում դիֆուզիոն փոխանցման երեւույթների հետ հանդիսանում է բջջի կենսագործունեությունն ապահովող հիմքը։
ԵՎ ակտիվտրանսպորտ. Պասիվ տրանսպորտը տեղի է ունենում առանց էներգիայի սպառման էլեկտրաքիմիական գրադիենտի երկայնքով: Պասիվները ներառում են դիֆուզիոն (պարզ և հեշտացված), օսմոզ, ֆիլտրացիա: Ակտիվ տրանսպորտը պահանջում է էներգիա և տեղի է ունենում կոնցենտրացիայի կամ էլեկտրական գրադիենտների դեմ:
Ակտիվ տրանսպորտ
Սա կոնցենտրացիայի կամ էլեկտրական գրադիենտներին հակառակ նյութերի տեղափոխումն է, որը տեղի է ունենում էներգիայի ծախսումով: Տարբերում են առաջնային ակտիվ տրանսպորտ, որը պահանջում է ATP էներգիա և երկրորդային (ATP-ի հաշվին մեմբրանի երկու կողմերում իոնային կոնցենտրացիայի գրադիենտների ստեղծում, և այդ գրադիենտների էներգիան օգտագործվում է փոխադրման համար):
Առաջնային ակտիվ տրանսպորտը լայնորեն կիրառվում է մարմնում: Այն մասնակցում է բջջային մեմբրանի ներքին և արտաքին կողմերի միջև էլեկտրական պոտենցիալ տարբերության ստեղծմանը: Ակտիվ տրանսպորտի օգնությամբ բջջի մեջտեղում և արտաբջջային հեղուկում ստեղծվում են Na +, K+, H+, SI «» և այլ իոնների տարբեր կոնցենտրացիաներ։
Ավելի լավ է ուսումնասիրված Na+ և K+ փոխադրումները՝ Na+, -K + -Hacoc։ Այս փոխադրումը տեղի է ունենում մոտ 100,000 մոլեկուլային քաշով գնդիկավոր սպիտակուցի մասնակցությամբ ներքին մակերեսըև արտաքին մակերեսի վրա երկու K+ կապող տեղամասեր: Սպիտակուցի ներքին մակերեսին նկատվում է ATPase-ի բարձր ակտիվություն։ ATP-ի հիդրոլիզի ընթացքում առաջացած էներգիան հանգեցնում է սպիտակուցի կոնֆորմացիոն փոփոխությունների և միաժամանակ բջջից հանվում են Na + 3 իոններ, իսկ այդպիսի պոմպի միջոցով ներմուծվում են երկու K+ իոններ Na+-ի բարձր կոնցենտրացիան առաջանում է արտաբջջային հեղուկում, իսկ K+–ի բարձր կոնցենտրացիան՝ բջջային հեղուկում։
IN ՎերջերսԻնտենսիվ ուսումնասիրվում են Ca2+ պոմպերը, որոնց շնորհիվ բջջում Ca2+-ի կոնցենտրացիան տասնյակ հազարավոր անգամ ցածր է, քան դրանից դուրս։ Բջջաթաղանթում և բջջի օրգանելներում կան Ca2+ պոմպեր (սարկոպլազմիկ ցանց, միտոքոնդրիա)։ Ca2+ պոմպերը գործում են նաև թաղանթներում առկա սպիտակուցի շնորհիվ: Այս սպիտակուցն ունի բարձր ATPase ակտիվություն:
Երկրորդային ակտիվ տրանսպորտ. Առաջնային ակտիվ տրանսպորտի շնորհիվ բջջից դուրս Na +-ի բարձր կոնցենտրացիա է ստեղծվում, պայմաններ են առաջանում Na +-ի բջջի մեջ տարածվելու համար, բայց Na+-ի հետ միասին կարող են ներթափանցել այլ նյութեր։ Այս տրանսպորտն ուղղված է մեկ ուղղությամբ և կոչվում է simport։ Հակառակ դեպքում, Na +-ի մուտքը խթանում է մեկ այլ նյութի ելքը բջջից, դրանք երկու հոսքեր են, որոնք ուղղված են տարբեր ուղղություններով.
Սիմպորտի օրինակ կարող է լինել գլյուկոզայի կամ ամինաթթուների տեղափոխումը Na +-ի հետ միասին: Փոխադրող սպիտակուցն ունի Na +-ի և գլյուկոզայի կամ ամինաթթուների միացման երկու տեղ: Հինգ տարբեր սպիտակուցներ են հայտնաբերվել, որոնք կապում են հինգ տեսակի ամինաթթուներ: Հայտնի են նաև սիմպորտի այլ տեսակներ՝ N+-ի տեղափոխումը բջիջի հետ միասին, K+ և Cl-ը բջիջից և այլն։
Գրեթե բոլոր բջիջներում գործում է հակապորտային մեխանիզմ՝ Na +-ը մտնում է բջիջ, իսկ Ca2+-ը դուրս է գալիս այն, կամ Na+-ը մտնում է բջիջ, իսկ H+-ը դուրս է գալիս դրանից։
Mg2 +, Fe2 +, HCO3- և շատ այլ նյութեր ակտիվորեն տեղափոխվում են թաղանթով:
Պինոցիտոզը ակտիվ տրանսպորտի տեսակներից մեկն է։ Այն կայանում է նրանում, որ որոշ մակրոմոլեկուլներ (հիմնականում սպիտակուցներ, որոնց մակրոմոլեկուլներն ունեն 100-200 նմ տրամագիծ) միանում են թաղանթային ընկալիչներին։ Այս ընկալիչները հատուկ են տարբեր սպիտակուցների համար: Նրանց կցումն ուղեկցվում է բջջի կծկվող սպիտակուցների՝ ակտինի և միոզինի ակտիվացմամբ, որոնք ձևավորում և փակում են խոռոչ այս արտաբջջային սպիտակուցով և արտաբջջային փոքր քանակությամբ հեղուկով։ Այս դեպքում ձևավորվում է պինոցիտոտիկ վեզիկուլ: Այն արտազատում է ֆերմենտներ, որոնք հիդրոլիզացնում են այս սպիտակուցը: Հիդրոլիզի արտադրանքը ներծծվում է բջիջների կողմից: Պինոցիտոզը պահանջում է ATP էներգիա և Ca2+-ի առկայությունը արտաբջջային միջավայրում:
Այսպիսով, կան բազմաթիվ տեսակի նյութերի տեղափոխում բջջային մեմբրաններով: Բջջի տարբեր կողմերում (գագաթային, բազալային, կողային թաղանթներում) կարող են առաջանալ տարբեր տեսակներտրանսպորտ. Դրա օրինակ կարող են լինել գործընթացները, որոնք տեղի են ունենում
Որոշ անօրգանական իոնների փոխանցումը տեղի է ունենում կոնցենտրացիայի գրադիենտի դեմ՝ տրանսպորտային ATPases (իոնային պոմպեր) մասնակցությամբ։ Բոլոր իոնային պոմպերը միաժամանակ ծառայում են որպես ավտոֆոսֆորիլացման և աուտոֆոսֆորիլացման ընդունակ ֆերմենտներ: ATPազները տարբերվում են իոնների առանձնահատկություններով, տեղափոխվող իոնների քանակով և փոխադրման ուղղությամբ։ ATPase-ի աշխատանքի արդյունքում տեղափոխված իոնները կուտակվում են մեմբրանի մի կողմում։ Մարդու բջիջների պլազմային թաղանթում ամենատարածվածներն են ստամոքսի լորձաթաղանթի Na + ,K + -ATPase, Ca 2+ -ATPase և H + ,K + , -ATPase:
Նա + , ՏՈ + -ATPase
Այս տրանսպորտային ֆերմենտը կատալիզացնում է ATP-ից կախված Na + և K + իոնների փոխադրումը պլազմային թաղանթով: Na + ,K + -ATPase-ը բաղկացած է α և β ենթամիավորներից; α - կատալիտիկ մեծ ենթաբաժին, β - փոքր ենթաբաժին (գլյուկոպրոտեին): Տրանսլոկազի ակտիվ ձևը տետրամեր է (αβ)2։
Na + ,K + -ATPase-ն պատասխանատու է բջիջում K + բարձր կոնցենտրացիայի և Na + ցածր կոնցենտրացիայի պահպանման համար: Քանի որ Na + ,K + -ATPase-ն դուրս է մղում երեք դրական լիցքավորված իոններ և պոմպեր է երկու մասի, բջջի ներսի մեմբրանի վրա հայտնվում է բացասական արժեք ունեցող էլեկտրական պոտենցիալ՝ արտաքին մակերեսի համեմատ:
Ք.ա 2+ -ATPase
«Հանգիստ» բջիջների ցիտոզոլում Ca 2+-ի կոնցենտրացիան կազմում է ~10"7 մոլ/լ, իսկ բջջից դուրս՝ ~2-10-3 մոլ/լ։ Կոնցենտրացիայի այս տարբերությունը պահպանվում է ակտիվ համակարգի կողմից։ կալցիումի իոնների տեղափոխում նրա հիմնական բաղադրիչներն են կալցիումի պոմպերը՝ Ca 2+ -ATPases և Na + ,Ca 2+ -փոխանակիչներ։
Ca 2+ -ATPase-ը տեղայնացված է ոչ միայն պլազմային թաղանթում, այլև ER թաղանթում։ Ֆերմենտը բաղկացած է տասը տրանսմեմբրանային տիրույթներից, որոնք տարածվում են բջջային թաղանթի վրա: Երկրորդ և երրորդ տիրույթների միջև կան ասպարտիկ թթվի մի քանի մնացորդներ, որոնք ներգրավված են կալցիումի կապակցման մեջ: Չորրորդ և հինգերորդ տիրույթների միջև ընկած հատվածն ունի ATP-ի կապի և ավտոֆոսֆորիլացման տեղ ասպարտիկ թթվի մնացորդի մոտ: Որոշ բջիջների պլազմային թաղանթների Ca 2+ -ATPa-ները կարգավորվում են կալմոդուլին սպիտակուցով։ Պլազմային մեմբրանի և ER-ի Ca 2+ -ATPa-երից յուրաքանչյուրը ներկայացված է մի քանի իզոֆորմներով:
Ca-ի ակտիվության խախտում 2+ -ATPases պաթոլոգիայում.Խափանման պատճառներից մեկը
Ca 2+ -ATPase - մեմբրանների լիպիդային պերօքսիդացման (LPO) ակտիվացում: Ֆոսֆոլիպիդներում և ֆերմենտի ակտիվ կենտրոնում գտնվող SH խմբերի ճարպաթթուների ացիլային մնացորդները ենթակա են օքսիդացման: Լիպիդային միջավայրի կառուցվածքի և ակտիվ կենտրոնի կառուցվածքի խախտումը հանգեցնում է ATPase-ի կոնֆորմացիայի փոփոխության, կալցիումի իոնների նկատմամբ մերձեցման կորստի և աուտոֆոսֆորիլացման ունակության: ATPase-ն դադարեցնում է կալցիումի իոնների դուրս մղումը բջջի ցիտոզոլից, ներբջջային կալցիումի կոնցենտրացիան մեծանում է, Ca 2+ մեծացնում է մկանների կծկումը, իսկ մկանային պատի տոնուսը մեծանում է, ինչը հանգեցնում է արյան ճնշման բարձրացման։ Ca 2+ -ATPase-ի ֆունկցիայի խանգարումը կարևոր դեր է խաղում աթերոսկլերոզի, քաղցկեղի և իմունային պաթոլոգիաների առաջացման գործում:
2. Երկրորդային ակտիվ տրանսպորտ
Որոշ լուծույթների տեղափոխումը կոնցենտրացիայի գրադիենտի նկատմամբ կախված է միաժամանակյա կամ հաջորդական փոխադրումից
մեկ այլ նյութ նույն ուղղությամբ համակենտրոնացման գրադիենտի երկայնքով (ակտիվ սիմպորտ)կամ հակառակը (ակտիվ հակապորտ):Մարդու բջիջներում կոնցենտրացիայի գրադիենտով տեղափոխվող իոնն ամենից հաճախ Na + է:
Տրանսպորտի այս տեսակի օրինակն է պլազմային մեմբրանի Na +, Ca 2+ փոխանակիչը (ակտիվ հակապորտ), նատրիումի իոնները կոնցենտրացիայի գրադիենտի երկայնքով տեղափոխվում են բջիջ, իսկ Ca 2+ իոնները՝ ընդդեմ կոնցենտրացիայի գրադիենտի, թողնում են բջիջը։
Ակտիվ սիմպորտի մեխանիզմի համաձայն՝ գլյուկոզան ներծծվում է աղիքային բջիջների կողմից, իսկ գլյուկոզան և ամինաթթուները՝ առաջնային մեզից երիկամների բջիջների կողմից:
մակրոմոլեկուլների և մասնիկների տեղափոխում մեմբրանի միջոցովԷՆԴՈՑԻՏՈԶ ԵՎ ԶԿՈՑԻՏՈԶ
Տրանսպորտային սպիտակուցներն ապահովում են փոքր բևեռային մոլեկուլների շարժումը բջջային թաղանթով, բայց նրանք չեն կարող տեղափոխել մակրոմոլեկուլներ, ինչպիսիք են սպիտակուցները, նուկլեինաթթուները, պոլիսախարիդները կամ նույնիսկ ավելի մեծ մասնիկները: Մեխանիզմները, որոնց միջոցով բջիջները կարող են վերցնել այդպիսի նյութերը կամ հեռացնել դրանք բջջից, տարբերվում են իոնների և բևեռային միացությունների տեղափոխման մեխանիզմներից։
Մեմբրանային տրանսպորտային սպիտակուցները մասնակցում են պլազմալեմայի միջոցով իոնների տեղափոխմանը։ Այս սպիտակուցները կարող են մեկ նյութ տեղափոխել մեկ ուղղությամբ (uniport) կամ մի քանի նյութեր միաժամանակ (symport), ինչպես նաև, մեկ նյութի ներմուծման հետ մեկտեղ, հեռացնել մյուսը բջիջից (antiport): Գլյուկոզան, օրինակ, Na+ իոնի հետ միասին կարող է ներթափանցել բջիջներ։ Իոնների տեղափոխումը կարող է տեղի ունենալ կոնցենտրացիայի գրադիենտի երկայնքով, այսինքն՝ պասիվ, առանց լրացուցիչ էներգիայի սպառման: Պասիվ փոխադրման դեպքում որոշ մեմբրանի փոխադրող սպիտակուցներ ձևավորում են մոլեկուլային բարդույթներ՝ ալիքներ, որոնցով լուծված մոլեկուլները անցնում են մեմբրանի միջով՝ պարզ դիֆուզիայի միջոցով կոնցենտրացիայի գրադիենտով։ Այս ալիքներից մի քանիսը մշտապես բաց են, մյուսները կարող են փակվել կամ բացվել՝ ի պատասխան ազդանշանային մոլեկուլների հետ կապվելու կամ ներբջջային իոնների կոնցենտրացիայի փոփոխության: Այլ դեպքերում հատուկ թաղանթային կրող սպիտակուցները ընտրողաբար կապվում են այս կամ այն իոնի հետ և տեղափոխում այն մեմբրանի միջով (հեշտացված դիֆուզիոն)։ Բջիջների ցիտոպլազմայում իոնների կոնցենտրացիան կտրուկ տարբերվում է ոչ միայն արտաքին միջավայրի կոնցենտրացիայից, այլև նույնիսկ արյան պլազմայից, որը լվանում է ավելի բարձր կենդանիների մարմնի բջիջները: Բջիջների ներսում և դրսում միավալենտ կատիոնների ընդհանուր կոնցենտրացիան գրեթե նույնն է (150 մՄ), իզոտոնիկ։ Բայց ցիտոպլազմայում K+-ի կոնցենտրացիան գրեթե 50 անգամ ավելի բարձր է, իսկ Na+-ն ավելի ցածր է, քան արյան պլազմայում, և այդ տարբերությունը պահպանվում է միայն կենդանի բջջում. ապա որոշ ժամանակ անց իոնային տարբերությունները երկու կողմերից պլազմային թաղանթը կվերանա: Պարզապես կարելի է բջիջները սառեցնել մինչև +2 o C, և որոշ ժամանակ անց մեմբրանի երկու կողմերում K+-ի և Na+-ի կոնցենտրացիաները կդառնան նույնը։ Երբ բջիջները տաքացվում են, այդ տարբերությունը վերականգնվում է։ Այս երեւույթը պայմանավորված է նրանով, որ բջիջներում կան թաղանթային սպիտակուցային կրիչներ, որոնք աշխատում են կոնցենտրացիայի գրադիենտի դեմ՝ միաժամանակ էներգիա ծախսելով ATP հիդրոլիզի շնորհիվ։ Նյութի փոխանցման այս տեսակը կոչվում է ակտիվ տրանսպորտ, և այն իրականացվում է սպիտակուցային իոնային պոմպերի միջոցով: Պլազմային թաղանթը պարունակում է երկու ենթաբաժնի մոլեկուլ (K+ + Na+)՝ պոմպ, որը նույնպես ATPase է։ Այս պոմպը մեկ ցիկլով դուրս է մղում 3 Na+ իոններ և 2 K+ իոններ մղում է բջիջ՝ ընդդեմ կոնցենտրացիայի գրադիենտի։ Այս դեպքում մեկ ATP մոլեկուլ ծախսվում է ATPase-ի ֆոսֆորիլացման վրա, որի արդյունքում Na+-ը բջջից տեղափոխվում է թաղանթով, իսկ K+-ը հնարավորություն է ստանում կապ հաստատել սպիտակուցի մոլեկուլի հետ, այնուհետև տեղափոխել բջիջ։ Թաղանթային պոմպերի միջոցով ակտիվ տեղափոխման արդյունքում բջջում կարգավորվում է նաև երկվալենտ կատիոնների՝ Mg 2+ և Ca + կոնցենտրացիան՝ նաև ATP-ի սպառմամբ։ Ակտիվ իոնային տրանսպորտի հետ միասին տարբեր շաքարներ, նուկլեոտիդներ և ամինաթթուներ թափանցում են պլազմային թաղանթ։ Այսպիսով, գլյուկոզայի ակտիվ փոխադրումը, որը միաժամանակ թափանցում է բջիջ պասիվ տեղափոխվող Na+ իոնի հոսքի հետ մեկտեղ, կախված կլինի (K+, Na+) պոմպի ակտիվությունից։ Եթե այս պոմպն արգելափակվի, ապա շուտով մեմբրանի երկու կողմերում Na+-ի կոնցենտրացիայի տարբերությունը կվերանա, մինչդեռ Na+-ի դիֆուզիան բջջի մեջ կնվազի, և միևնույն ժամանակ գլյուկոզայի հոսքը բջիջ կդադարի։ Հենց վերականգնվում է (K+ + Na+)-ATPase-ի աշխատանքը և ստեղծվում է իոնների կոնցենտրացիայի տարբերություն, Na+-ի ցրված հոսքը անմիջապես մեծանում է և, միաժամանակ, գլյուկոզայի տեղափոխումը։ Սրա նման
իրականացվում է ամինաթթուների տեղափոխում, որոնք մեմբրանի միջով տեղափոխվում են հատուկ կրող սպիտակուցներով, որոնք գործում են որպես սիմպորտային համակարգեր՝ միաժամանակ տեղափոխելով իոններ։ Բակտերիաների բջիջներում շաքարների և ամինաթթուների ակտիվ տեղափոխումը պայմանավորված է ջրածնի իոնների գրադիենտով: Ցածր մոլեկուլային միացությունների պասիվ կամ ակտիվ փոխադրմանը հատուկ թաղանթային սպիտակուցների բուն մասնակցությունը ցույց է տալիս այս գործընթացի բարձր յուրահատկությունը։ Նույնիսկ պասիվ իոնների փոխադրման դեպքում սպիտակուցները «ճանաչում» են տվյալ իոնը, փոխազդում են նրա հետ, հատուկ կապվում, փոխում են դրանց կոնֆորմացիան և գործառույթը։ Հետևաբար, նույնիսկ պարզ նյութերի տեղափոխման օրինակում թաղանթները հանդես են գալիս որպես անալիզատորներ և ընկալիչներ։ Մեմբրանի ընկալիչի ֆունկցիան հատկապես ակնհայտ է, երբ բջիջը կլանում է կենսապոլիմերները։
Միջբջջային շփումներ.
Բազմաբջիջ օրգանիզմներում միջբջջային փոխազդեցությունների շնորհիվ առաջանում են բարդ բջջային հավաքներ, որոնց պահպանումն իրականացվում է տարբեր եղանակներով։ Բողբոջային, սաղմնային հյուսվածքներում, հատկապես զարգացման վաղ փուլերում, բջիջները մնում են միմյանց հետ կապված՝ շնորհիվ իրենց մակերեսների միմյանց կպչելու ունակության։ Այս գույքը կպչունությունԲջիջների (միացում, կպչունություն) կարելի է որոշել դրանց մակերեսի հատկություններով, որոնք հատուկ փոխազդում են միմյանց հետ։ Երբեմն, հատկապես միաշերտ էպիթելիայում, հարևան բջիջների պլազմային թաղանթները ձևավորում են բազմաթիվ ինվագինացիաներ, որոնք հիշեցնում են ատաղձագործի կարը։ Սա լրացուցիչ ուժ է ստեղծում միջբջջային կապի համար: Նման պարզ սոսինձային (բայց հատուկ) միացումից բացի, կան մի շարք հատուկ միջբջջային կառուցվածքներ, կոնտակտներ կամ կապեր, որոնք կատարում են հատուկ գործառույթներ: Սրանք կողպման, խարսխման և կապի միացումներ են: Փական, թե խիտ,կապը բնորոշ է միաշերտ էպիթելիային։ Սա այն գոտին է, որտեղ երկու պլազմային թաղանթների արտաքին շերտերը հնարավորինս մոտ են: Այս շփման ժամանակ մեմբրանի եռաշերտ կառուցվածքը հաճախ տեսանելի է. երկու թաղանթների արտաքին օսմիոֆիլ շերտերը կարծես միաձուլվել են մեկ ընդհանուր շերտի մեջ՝ 2-3 նմ հաստությամբ: Օգտագործելով պինդ շփման գոտում պլազմային մեմբրանի ընդմիջումների հարթ պատրաստուկներ, սառեցման և մանրացման մեթոդի կիրառմամբ, պարզվել է, որ թաղանթների շփման կետերը գնդիկներ են (ամենայն հավանականությամբ, պլազմային մեմբրանի հատուկ ինտեգրալ սպիտակուցներ) շարքեր. Գնդիկների նման շարքերը կամ շերտերը կարող են հատվել այնպես, որ վանդակավոր կամ ցանց ձևավորեն չիպի մակերեսին։ Վերջին դեպքում պինդ կոնտակտը կազմում է պլազմային թաղանթների միաձուլման շարունակական գոտի՝ բջիջը շրջապատելով նրա գագաթային (վերին, աղիքային լույսի մեջ նայելով) մասում։ Այսպիսով, շերտի յուրաքանչյուր բջիջ, կարծես, շրջապատված է այս շփման ժապավենով: Հատուկ բծերի դեպքում նման կառույցները կարելի է տեսնել նաև լուսային մանրադիտակի տակ։ Նրանք ստացել են անվանման վերջի թիթեղները մորֆոլոգներից։ Այս դեպքում փակվող ամուր հանգույցի դերը միայն բջիջները միմյանց մեխանիկորեն միացնելը չէ: Այս կոնտակտային տարածքը վատ թափանցելի է մակրոմոլեկուլների և իոնների համար, և այդպիսով այն փակում և արգելափակում է միջբջջային խոռոչները՝ մեկուսացնելով դրանք (և դրանց հետ միասին մարմնի ներքին միջավայրը) արտաքին միջավայրից (այս դեպքում՝ աղիքային լույսը): Չնայած բոլոր ամուր հանգույցները խոչընդոտներ են մակրոմոլեկուլների համար, նրանց թափանցելիությունը փոքր մոլեկուլների նկատմամբ տարբեր էպիթելիաների միջև: խարսխում (միացում)կապերը կամ կոնտակտները կոչվում են այսպես, քանի որ դրանք ոչ միայն միացնում են հարևան բջիջների պլազմային թաղանթները, այլև հաղորդակցվում են ցիտոկմախքի ֆիբրիլային տարրերի հետ: Այս տեսակի միացությունը բնութագրվում է երկու տեսակի սպիտակուցների առկայությամբ. Դրանցից մեկը ներկայացված է տրանսմեմբրանային կապող (կապող) սպիտակուցներով, որոնք ներգրավված են կա՛մ միջբջջային կապի մեջ, կա՛մ պլազմային մեմբրանի միացման մեջ արտաբջջային մատրիցայի բաղադրիչների հետ (էպիթելի նկուղային թաղանթ, շարակցական հյուսվածքի արտաբջջային կառուցվածքային սպիտակուցներ): Երկրորդ տեսակը ներառում է ներբջջային սպիտակուցներ, որոնք կապում կամ խարսխում են նման շփման թաղանթային տարրերը ցիտոկմախքի ցիտոպլազմիկ մանրաթելերի հետ։ Միջբջջային կետային կպչուն հանգույցներ հայտնաբերվում են շատ ոչ էպիթելային հյուսվածքներում, սակայն սոսնձային հանգույցների կառուցվածքն ավելի հստակ է նկարագրված: ժապավեններ, կամ գոտիներ, միաշերտ էպիթելիայում։ Այս կառուցվածքը շրջապատում է էպիթելի բջջի ողջ պարագիծը, նման է այն, ինչ տեղի է ունենում ամուր միացման դեպքում: Ամենից հաճախ նման գոտին կամ ժապավենը գտնվում է ամուր կապի տակ: Այս վայրում պլազմային թաղանթները մոտեցվում են միմյանց, և նույնիսկ մի փոքր հեռու են միմյանցից 25 - 30 նմ հեռավորության վրա, և նրանց միջև տեսանելի է աճող խտության գոտի: Սրանք ոչ այլ ինչ են, քան տրանսմեմբրանային գլիկոպրոտեինների փոխազդեցության վայրեր, որոնք Ca++ իոնների մասնակցությամբ հատուկ կպչում են միմյանց և ապահովում են երկու հարևան բջիջների թաղանթների մեխանիկական կապը։ Կապիչ սպիտակուցները պատկանում են կադերիններին՝ ընկալիչ սպիտակուցներ, որոնք ապահովում են բջիջների կողմից համասեռ թաղանթների հատուկ ճանաչումը: Գլիկոպրոտեինային շերտի ոչնչացումը հանգեցնում է առանձին բջիջների տարանջատմանը և էպիթելի շերտի ոչնչացմանը։ Ցիտոպլազմային կողմում, թաղանթի մոտ, տեսանելի է խիտ նյութի կուտակում, որին հարում է պլազմային թաղանթի երկայնքով ընկած բարակ (6 - 7 նմ) թելերի շերտը ամբողջ պարագծի երկայնքով ձգվող կապոցի տեսքով։ բջիջ. Նիհար թելերը դասակարգվում են որպես ակտինի մանրաթելեր; նրանք կապվում են պլազմային թաղանթին սպիտակուցի վինկուլինի միջոցով, որը կազմում է խիտ կողային թաղանթ: Ժապավենի միացման ֆունկցիոնալ նշանակությունը ոչ միայն բջիջների միմյանց մեխանիկական կպչունության մեջ է. երբ ժապավենի մեջ ակտինի թելերը կծկվեն, բջջի ձևը կարող է փոխվել: Կիզակետային կոնտակտներ կամ կալանքային թիթեղներ, հայտնաբերվել են բազմաթիվ բջիջներում և հատկապես լավ ուսումնասիրված են ֆիբրոբլաստներում։ Դրանք կառուցված են ընդհանուր հատակագծի համաձայն՝ կպչուն ժապավեններով, բայց արտահայտվում են փոքր հատվածների՝ պլազմալեմայի վրա սալիկների տեսքով։ Այս դեպքում տրանսմեմբրանային կապող սպիտակուցները հատուկ կապվում են արտաբջջային մատրիցային սպիտակուցների հետ, ինչպիսին է ֆիբրոնեկտինը: Ցիտոպլազմային կողմում այս նույն գլիկոպրոտեինները կապված են մոտ թաղանթային սպիտակուցների հետ, որոնք ներառում են վինկուլինը, որն իր հերթին կապված է ակտինի թելերի փաթեթի հետ: Կիզակետային կոնտակտների ֆունկցիոնալ նշանակությունը և՛ բջիջներն արտաբջջային կառույցներին խարսխելու, և՛ բջիջների շարժման հնարավորություն տվող մեխանիզմի ապահովումն է: Դեզմոսոմներ, որոնք նման են սալիկների կամ կոճակների, նաև բջիջները միացնում են միմյանց։ Միջբջջային տարածությունում այստեղ տեսանելի է նաև խիտ շերտ, որը ներկայացված է փոխազդող ինտեգրալ թաղանթային գլիկոպրոտեիններով՝ դեզմոգլեյններով, որոնք նույնպես, կախված Ca++ իոններից, կպչում են բջիջները միմյանց։ Ցիտոպլազմային կողմում պլազմալեմային կից է դեզմոպլակին սպիտակուցի շերտը, որի հետ կապված են ցիտոկմախքի միջանկյալ թելերը։ Դեզմոսոմներն առավել հաճախ հանդիպում են էպիթելիայում, որի դեպքում միջանկյալ թելերը պարունակում են կերատիններ։ Սրտի մկանային բջիջները՝ կարդիոմիոցիտները, պարունակում են դեզմինի մանրաթելեր՝ որպես դեզմոսոմների մաս։ Անոթային էնթելիումում դեսմոսոմները պարունակում են վիմենտինի միջանկյալ թելեր։ Հեմիդեսմոսոմները կառուցվածքով նման են դեզմոսոմներին, բայց ներկայացնում են բջիջների կապը միջբջջային կառուցվածքների հետ։ Դեսմոսոմների և հեմիդեսմոսոմների ֆունկցիոնալ դերը զուտ մեխանիկական է. նրանք կպչում են բջիջները միմյանց և հիմքում ընկած արտաբջջային մատրիցին: Ի տարբերություն ամուր շփման, բոլոր տեսակները կպչուն կոնտակտներթափանցելի է ջրային լուծույթներև որևէ դեր չեն խաղում դիֆուզիայի սահմանափակման գործում: Սլոտի կոնտակտներհամարվում են բջիջների հաղորդակցման հանգույցներ: Այս կառույցները ներգրավված են ուղղակի փոխանցման մեջ քիմիական նյութերբջիջից բջիջ: Շփման այս տեսակը բնութագրվում է երկու հարևան բջիջների պլազմային թաղանթների 2-3 նմ հեռավորության վրա միացնելով: Օգտագործելով սառեցման մեթոդը: Պարզվեց, որ ճեղքված թաղանթների վրա բացերի կոնտակտների գոտիները (0,5-ից մինչև 5 մկմ չափի) կետավոր են 7-8 նմ տրամագծով մասնիկներով, որոնք դասավորված են վեցանկյուն՝ 8-10 նմ ժամանակահատվածով և ունեն մոտ 2 հոր լայնությամբ ալիք։ կենտրոնը։ Այս մասնիկները կոչվում են կոնքսոններ։ Բաց միացման գոտիներում կարող են լինել 10-ից 20-ից մինչև մի քանի հազար կապեր՝ կախված բջիջների ֆունկցիոնալ բնութագրերից: Նախապատրաստականորեն մեկուսացվել են կոնքսոնները: Դրանք կազմված են կոնեկտինի՝ սպիտակուցի վեց ենթամիավորներից։ Համակցվելով միմյանց հետ՝ կոնեկտինները կազմում են գլանաձև ագրեգատ՝ միացում, որի կենտրոնում կա ալիք։ Առանձին կապակցիչներ տեղադրվում են պլազմային թաղանթում այնպես, որ նրանք թափանցում են այն անմիջապես միջով: Բջջի պլազմային թաղանթի վրա գտնվող մեկ կապակցումը ճշգրիտ հակադրվում է հարակից բջջի պլազմային թաղանթի վրա գտնվող միացմանը, այնպես որ երկու միացումների ալիքները կազմում են մեկ միավոր: Կապակցումները կատարում են ուղիղ միջբջջային ուղիների դեր, որոնց միջոցով իոնները և ցածր մոլեկուլային քաշ ունեցող նյութերը կարող են ցրվել բջջից բջիջ: Կապակցումները կարող են փակվել՝ փոխելով ներքին ալիքի տրամագիծը և դրանով իսկ մասնակցել բջիջների միջև մոլեկուլների փոխադրման կարգավորմանը։ Ոչ սպիտակուցները, ոչ էլ նուկլեինաթթուները չեն կարող անցնել բացերի հանգույցներով: Ցածր մոլեկուլային քաշի միացություններ անցնելու բացերի միացումների կարողությունը ընկած է էլեկտրական իմպուլսի (գրգռման ալիքի) արագ փոխանցման հիմքում բջիջից բջիջ առանց նյարդային հաղորդիչի մասնակցության: Սինապտիկ կոնտակտ (սինապսներ). Սինապսները երկու բջիջների միջև շփման տարածքներ են, որոնք մասնագիտացված են մի տարրից մյուսը գրգռման կամ արգելակման միակողմանի փոխանցման համար: Շփման այս տեսակը բնորոշ է նյարդային հյուսվածքին և առաջանում է ինչպես երկու նեյրոնների, այնպես էլ նեյրոնների և մի այլ տարրի՝ ընկալիչի կամ էֆեկտորի միջև: Սինապտիկ շփման օրինակ է նաև նյարդամկանային վերջավորությունը։ Ինտերնեյրոնային սինապսները սովորաբար ունենում են տանձաձև ընդարձակման (տախտակների) տեսք։ Սինապտիկ թիթեղները կարող են շփվել ինչպես մեկ այլ նեյրոնի մարմնի, այնպես էլ դրա գործընթացների հետ: Նյարդային բջիջների (աքսոնների) ծայրամասային պրոցեսները հատուկ շփումներ են ստեղծում էֆեկտոր բջիջների (մկանային կամ գեղձային) կամ ընկալիչ բջիջների հետ։ Հետևաբար, սինապսը մասնագիտացված կառույց է, որը ձևավորվում է երկու բջիջների շրջանների միջև (ճիշտ այնպես, ինչպես դեզմոսոմը): Սինապտիկ շփումների վայրերում բջջային թաղանթները բաժանված են միջբջջային տարածությամբ՝ մոտ 20-30 նմ լայնությամբ սինապտիկ ճեղքվածքով: Հաճախ բացվածքի լուսանցքում տեսանելի է թաղանթներին ուղղահայաց տեղակայված նուրբ թելքավոր նյութը։ Մի բջջի թաղանթը, որը գրգռում է փոխանցում սինապտիկ շփման տարածքում, կոչվում է նախասինապտիկ, մյուս բջիջի թաղանթը, որն ընդունում է իմպուլսը, կոչվում է հետսինապտիկ: Նախասինապտիկ մեմբրանի մոտ հայտնաբերվում է մեծ գումարփոքր վակուոլներ - սինապտիկ վեզիկուլներ, որոնք լցված են հաղորդիչներով: Սինապտիկ վեզիկուլների պարունակությունը նյարդային ազդակի անցման ժամանակ էկզոցիտոզով ազատվում է սինապտիկ ճեղքվածքի մեջ։ Հետսինապտիկ թաղանթը հաճախ ավելի հաստ է թվում, քան սովորական թաղանթները՝ ցիտոպլազմային կողմում իր մոտ բազմաթիվ բարակ մանրաթելերի կուտակման պատճառով: Պլազմոդեզմատա.Միջբջջային հաղորդակցության այս տեսակը հանդիպում է բույսերում: Plasmodesmata-ն բարակ խողովակային ցիտոպլազմային ալիքներ են, որոնք միացնում են երկու հարակից բջիջները: Այս ալիքների տրամագիծը սովորաբար 20 - 40 նմ է: Այս ալիքները սահմանափակող թաղանթն ուղղակիորեն անցնում է հարևան բջիջների պլազմային թաղանթների մեջ։ Պլազմոդեզմատան անցնում է բջիջները բաժանող բջջային պատի միջով: Հարևան բջիջների էնդոպլազմիկ ցանցի ցիստեռնները միացնող թաղանթավոր խողովակային տարրերը կարող են ներթափանցել պլազմոդեզմատայի ներսում: Պլազմոդեզմատաները ձևավորվում են բաժանման ժամանակ, երբ առաջնային Բջջային թաղանթ. Նոր բաժանված բջիջներում պլազմոդեզմատների թիվը կարող է շատ մեծ լինել (մինչև 1000 մեկ բջջում)։ Բջիջների ծերացման հետ մեկտեղ դրանց թիվը նվազում է պատերի հաստության մեծացման պատճառով ճեղքվածքների պատճառով: Լիպիդային կաթիլները կարող են շարժվել պլազմոդեզմատայի երկայնքով: Պլազմոդեզմատայի միջոցով բջիջները վարակվում են բույսերի վիրուսներով։
Ակտիվ տրանսպորտ- սա նյութի տեղափոխումն է ավելի ցածր էլեկտրաքիմիական ներուժ ունեցող վայրերից ավելի մեծ արժեք ունեցող վայրեր:
Ակտիվ փոխադրումը թաղանթում ուղեկցվում է Գիբսի էներգիայի ավելացմամբ, այն չի կարող տեղի ունենալ ինքնաբերաբար, բայց միայն ադենոզին եռաֆոսֆորական թթվի (ATP) հիդրոլիզի գործընթացի հետ միասին, այսինքն՝ բարձր էներգիայի ծախսման պատճառով. ATP-ի էներգետիկ պարտատոմսեր.
Նյութերի ակտիվ տեղափոխումը կենսաբանական թաղանթներով մեծ նշանակություն ունի: Ակտիվ տրանսպորտի շնորհիվ մարմնում ստեղծվում են կոնցենտրացիայի գրադիենտներ, էլեկտրական պոտենցիալ գրադիենտներ, ճնշման գրադիենտներ և այլն՝ աջակցելով կյանքի գործընթացներին, այսինքն. թերմոդինամիկայի տեսանկյունից ակտիվ փոխանցումը օրգանիզմը պահում է ոչ հավասարակշռված վիճակում և պահպանում կյանքը։
Կենսաբանական թաղանթներով նյութերի ակտիվ փոխադրման առկայությունը առաջին անգամ ապացուցվել է Ուսինգի (1949) փորձերում՝ օգտագործելով գորտի մաշկի միջով նատրիումի իոնների փոխանցման օրինակը (նկ. 12):
Բրինձ. 12.Ուսինգի փորձերի սխեման (A - ամպաչափ, V - վոլտմետր, B - մարտկոց, P - պոտենցիոմետր)
Ուսինգի փորձարարական խցիկը, որը լցված էր սովորական Ռինգերի լուծույթով, բաժանվեց երկու մասի՝ թարմ մեկուսացված գորտի կաշվով։ Նկ. 12, ձախում՝ մաշկի արտաքին լորձաթաղանթի մակերեսը, աջում՝ ներքին շիճուկը։ Դիտարկվել են նատրիումի իոնների հոսքեր գորտի մաշկի միջով՝ ձախից աջ արտաքինից դեպի ներքին մակերես և աջից ձախ՝ ներքինից դեպի արտաքին մակերես։
Թեորելի հավասարումից, որը նկարագրում է պասիվ տրանսպորտը, հետևում է Ussing-Theorell հավասարումըպասիվ տրանսպորտի դեպքում այս հոսքերի հարաբերակցության համար.
J m,in /j m,nar = (With out /With in)×e ZF j / RT
Ռինգերի լուծույթը բաժանող գորտի մաշկի վրա առաջանում է պոտենցիալ տարբերություն (j in - j nar) - մաշկի ներքին կողմը դրական ներուժ ունի արտաքինի նկատմամբ։ Ussing ինստալացիան (նկ. 12) ունեցել է լարման փոխհատուցման միավոր, որի օգնությամբ գորտի մաշկի վրա պոտենցիալ տարբերությունը զրոյացվել է, որը կառավարվում է վոլտմետրով։ Իոնների նույն կոնցենտրացիան պահպանվել է արտաքին և ներքին կողմերում՝ C out = C in:
Այս պայմաններում, եթե նատրիումի փոխանցումը գորտի մաշկի միջով որոշվում էր միայն պասիվ փոխադրմամբ, ապա Ussing-Theorell հավասարման համաձայն, j m, in և j m, nar հոսքերը հավասար էին միմյանց. j m, in =: ժ մ, նար
Մեմբրանի միջով ընդհանուր հոսքը կլինի զրո:
Ամպերաչափի միջոցով պարզվել է, որ փորձարարական պայմաններում (էլեկտրական ներուժի և կոնցենտրացիայի գրադիենտների բացակայություն) հոսում է գորտի մաշկի միջով։ էլեկտրաէներգիաԵս, հետևաբար կա լիցքավորված մասնիկների միակողմանի փոխանցում: Հաստատվել է, որ հոսանք մաշկի միջով անցնում է արտաքին միջավայրից դեպի ներքին միջավայր։
Փորձարարական տվյալները անհերքելիորեն ցույց են տվել, որ նատրիումի իոնների տեղափոխումը գորտի մաշկի միջով չի ենթարկվում պասիվ փոխադրման հավասարմանը: Հետեւաբար, ակտիվ փոխանցում է տեղի ունենում:
Էլեկտրական իոնային պոմպեր
Ժամանակակից հասկացությունների համաձայն, կենսաբանական թաղանթները պարունակում են իոնային պոմպեր,աշխատում է ATP հիդրոլիզի ազատ էներգիայի շնորհիվ, - հատուկ համակարգերինտեգրալ սպիտակուցներ (փոխադրող ATPases):
Ներկայումս հայտնի են երեք տեսակի էլեկտրագենային իոնային պոմպեր, որոնք ակտիվորեն տեղափոխում են իոններ թաղանթով (նկ. 13):
Տրանսպորտային ATPase-ներով իոնների փոխանցումը տեղի է ունենում տրանսպորտային գործընթացների հետ զուգակցման շնորհիվ քիմիական ռեակցիաներ, բջջային նյութափոխանակության էներգիայի շնորհիվ։
Երբ K + -Na + -ATPase-ն գործում է, յուրաքանչյուր ATP մոլեկուլի հիդրոլիզի ժամանակ թողարկված էներգիայի շնորհիվ կալիումի երկու իոն տեղափոխվում են բջիջ, իսկ երեք նատրիումի իոնները միաժամանակ դուրս են մղվում բջջից: Սա բջջում առաջացնում է կալիումի իոնների կոնցենտրացիայի ավելացում՝ համեմատած միջբջջային միջավայրի հետ, և նատրիումի կոնցենտրացիայի նվազում, ինչը մեծ ֆիզիոլոգիական նշանակություն ունի:
ATP հիդրոլիզի էներգիայի շնորհիվ երկու կալցիումի իոններ տեղափոխվում են Ca 2+ -ATPase, իսկ երկու պրոտոն՝ H + - պոմպ։
Նկ.13. Իոնային պոմպերի տեսակները. ա) K + -Na + - ATPase ցիտոպլազմային թաղանթներում.
(K + -Na + -pump); բ) - Ca 2+ -ATPase (Ca 2+ -pump); գ) - H + -ATPase միտոքոնդրիումների և քլորոպլաստների էներգիայի միացման մեմբրաններում (H + - պոմպ կամ պրոտոնային պոմպ)
Իոնային ATPase-ների գործողության մոլեկուլային մեխանիզմը լիովին պարզ չէ: Այնուամենայնիվ, կարելի է հետևել այս բարդ ֆերմենտային գործընթացի հիմնական փուլերին։ K + -Na + -ATPase-ի դեպքում կան իոնների փոխանցման յոթ փուլեր, որոնք կապված են ATP հիդրոլիզի հետ:
Դիագրամը ցույց է տալիս, որ ֆերմենտի հիմնական փուլերն են.
1) մեմբրանի ներքին մակերեսի վրա ATP-ով ֆերմենտային համալիրի ձևավորում (այս ռեակցիան ակտիվանում է մագնեզիումի իոններով).
2) երեք նատրիումի իոնների միացում համալիրի կողմից.
3) ֆերմենտի ֆոսֆորիլացում ադենոզին դիֆոսֆատի առաջացմամբ.
4) մեմբրանի ներսում ֆերմենտի պտույտ (flip-flop).
5) նատրիումի իոնափոխանակության ռեակցիան կալիումին, որը տեղի է ունենում մեմբրանի արտաքին մակերեսին.
6) ֆերմենտային համալիրի հակադարձ պտույտ՝ կալիումի իոնների բջիջ տեղափոխմամբ.
7) ֆերմենտի վերադարձը իր սկզբնական վիճակին՝ կալիումի իոնների և անօրգանական ֆոսֆատի (P) արտազատմամբ։
Այսպիսով, ամբողջական ցիկլի ընթացքում բջջից դուրս է գալիս նատրիումի երեք իոն, ցիտոպլազմը հարստանում է կալիումի երկու իոններով, և տեղի է ունենում մեկ ATP մոլեկուլի հիդրոլիզ։
Երկրորդային ակտիվ իոնային տրանսպորտ.
Բացի վերը քննարկված իոնային պոմպերից, հայտնի են նմանատիպ համակարգեր, որոնցում նյութերի կուտակումը կապված չէ ATP հիդրոլիզի հետ, այլ ռեդոքս ֆերմենտների կամ ֆոտոսինթեզի աշխատանքի հետ: Նյութերի տեղափոխումն այս դեպքում երկրորդական է՝ միջնորդավորված մեմբրանի ներուժով և/կամ իոնային կոնցենտրացիայի գրադիենտով՝ թաղանթում հատուկ կրիչների առկայության դեպքում: Տրանսպորտային այս մեխանիզմը կոչվում է երկրորդական ակտիվ տրանսպորտ: Այս մեխանիզմը առավել մանրամասն դիտարկվել է Պիտեր Միտչելի կողմից (1966 թ.) օքսիդատիվ ֆոսֆորիլացման քիմիոսմոտիկ տեսության մեջ: Կենդանի բջիջների պլազմայում և ենթաբջջային թաղանթներում հնարավոր է առաջնային և երկրորդային ակտիվ տրանսպորտի միաժամանակյա գործունեությունը: Օրինակ՝ միտոքոնդրիաների ներքին թաղանթն է։ Դրանում ATPase-ի արգելակումը մասնիկին չի զրկում երկրորդական ակտիվ տրանսպորտի պատճառով նյութեր կուտակելու ունակությունից։ Կուտակման այս մեթոդը հատկապես կարևոր է այն մետաբոլիտների համար, որոնց համար պոմպեր չկան (շաքարներ, ամինաթթուներ):
Ներկայումս բավական խորությամբ ուսումնասիրված են երկրորդային ակտիվ տրանսպորտի երեք սխեմաներ։ Դիտարկենք միավալենտ իոնների փոխադրումը կրող մոլեկուլների մասնակցությամբ։ Սա ենթադրում է, որ փոխադրողը բեռնված կամ բեռնաթափված վիճակում հավասարապես լավ հատում է թաղանթը: Էներգիայի աղբյուրը իոններից մեկի մեմբրանի ներուժն է և/կամ կոնցենտրացիայի գրադիենտը։ Շղթաները ներկայացված են Նկար 14-ում: Կոնկրետ կրիչի հետ համալիրում իոնի միակողմանի փոխանցումը կոչվում է uniport . Այս դեպքում լիցքը մեմբրանի վրայով փոխանցվում է կամ կոմպլեքսով, եթե կրող մոլեկուլը էլեկտրականորեն չեզոք է, կամ դատարկ կրիչով, եթե փոխանցումն ապահովվում է լիցքավորված կրիչով։ Փոխանցման արդյունքը կլինի իոնների կուտակումը մեմբրանի ներուժի նվազման պատճառով։ Այս ազդեցությունը նկատվում է, երբ կալիումի իոնները կուտակվում են էներգիա ունեցող միտոքոնդրիումներում վալինոմիցինի առկայության դեպքում:
Մեկ կրող մոլեկուլ պարունակող իոնների հակադարձ փոխանցումը կոչվում է հակապորտ . Ենթադրվում է, որ փոխադրող մոլեկուլը փոխանցում է յուրաքանչյուր իոնով ուժեղ բարդույթ։ Փոխանցումը տեղի է ունենում երկու փուլով՝ նախ մեկ իոնը ձախից աջ անցնում է թաղանթով, ապա երկրորդ իոնը՝ հակառակ ուղղությամբ։ Մեմբրանի ներուժը չի փոխվում: Ո՞րն է այս գործընթացի շարժիչ ուժը: Ակնհայտ է, որ փոխանցված իոններից մեկի կոնցենտրացիաների տարբերությունը: Եթե ի սկզբանե տարբերություն չի եղել երկրորդ իոնի կոնցենտրացիայի մեջ, ապա փոխանցման արդյունքը կլինի երկրորդ իոնի կուտակումը առաջինի կոնցենտրացիաների տարբերության նվազման պատճառով։ Հակապորտի դասական օրինակ է կալիումի և ջրածնի իոնների փոխանցումը բջջաթաղանթով՝ հակաբիոտիկ մոլեկուլ նիգերիցինի մասնակցությամբ:
Իոնների համատեղ միակողմանի փոխադրումը, որը ներառում է երկու տեղամասային փոխադրիչ, կոչվում է սիմպորտ . Ենթադրվում է, որ թաղանթը կարող է պարունակել երկու էլեկտրականորեն չեզոք մասնիկներ՝ կատիոնով և անիոնով կոմպլեքսավորված կրող և դատարկ կրող։ Քանի որ մեմբրանի պոտենցիալը չի փոխվում փոխանցման նման սխեմայի դեպքում, փոխանցումը կարող է առաջանալ իոններից մեկի կոնցենտրացիաների տարբերությամբ: Ենթադրվում է, որ սիմպորտի սխեման օգտագործվում է բջիջներում ամինաթթուների կուտակման համար: Կալիում-նատրիումային պոմպը (նկ. 13) ստեղծում է նատրիումի իոնների սկզբնական կոնցենտրացիայի գրադիենտ, որոնք այնուհետև, ըստ սիմպորտի սխեմայի, նպաստում են ամինաթթուների կուտակմանը։ Սիմպորտի սխեմայից հետևում է, որ այս գործընթացը պետք է ուղեկցվի օսմոտիկ հավասարակշռության զգալի տեղաշարժով, քանի որ մեկ ցիկլում երկու մասնիկ տեղափոխվում են մեմբրանի միջով նույն ուղղությամբ:
Նկար 14. Երկրորդային ակտիվ իոնային տրանսպորտի հիմնական սխեմաներ
Կյանքի ընթացքում բջիջների սահմանները հատում են տարբեր նյութեր, որոնց հոսքերը արդյունավետորեն կարգավորվում են։ Այս խնդիրն իրականացվում է բջջային թաղանթով, որի մեջ ներկառուցված են տրանսպորտային համակարգեր, ներառյալ իոնային պոմպեր, կրող մոլեկուլների համակարգ և բարձր ընտրողական իոնային ալիքներ:
Առաջին հայացքից փոխանցման համակարգերի նման առատությունը անհարկի է թվում, քանի որ միայն իոնային պոմպերի շահագործումը հնարավորություն է տալիս ապահովել կենսաբանական տրանսպորտի բնորոշ հատկանիշները. Այնուամենայնիվ, պարադոքսն այն է, որ կարգավորվող հոսքերի քանակը անսահման մեծ է, մինչդեռ կան ընդամենը երեք պոմպեր: Այս դեպքում առանձնահատուկ նշանակություն են ստանում իոնային կոնյուգացիայի մեխանիզմները, որոնք կոչվում են երկրորդական ակտիվ տրանսպորտ, որոնցում կարևոր դեր են խաղում դիֆուզիոն պրոցեսները։ Այսպիսով, նյութերի ակտիվ փոխադրման համադրությունը բջջաթաղանթում դիֆուզիոն փոխանցման երեւույթների հետ ապահովում է բջջի կենսագործունեությունը։